Lesson 6:
Lesson 6:
 Life's Origins
Life's Origins
|
6.6
Lateral Gene Transfers -
Crossing the Species Border
From the foregoing it is already clear that the concept of "species"
is of limited use in elucidating the origin of life and its evolution during the
early stages of history. The term species refers to interbreeding populations who
produce fertile offspring, or to potentially interbreeding populations (if they were at the
same place at the same time). It is a concept well suited for the classification of
domesticated plants and animals and all the other life forms which have similar life
histories. It is good to remember that the species concept originated in the 18th
century, when species were still considered immutable and bacteria
were unknown.
When it became clear that species are not immutable, the concept became fuzzy.
Genetic changes accumulate over time. Eventually, once-related populations
have evolved into separate and distinct species. But to draw the line where one species
ends and another begins in such a series is an arbitrary exercise.
Fortunately, evolution proceeds sometimes very slowly and sometimes
quite rapidly, based on the changing morphology seen in fossil
lineages, which makes it easier to draw a line. (This circumstance
has been celebrated by the impressive label "punctuated equilibrium".)
The main feature of a species is that its populations exchange genetic material, and such
exchange does not occur outside of the group that defines the
species. Now it seems that not only do we consist of modified
colonies of ancient types of bacteria, but we also carry at least
some genes from certain bacteria, short programs that have inserted
themselves into our replication code while we were evolving as
species. Apparently, evolution does not strictly proceed within
well-defined lineages. Instead "genetic noise" is penetrating the species barrier,
introducing an additional component of variability, in addition to internal mutation.
Of course, we are not special in this, it is happening to all the branches in the tree of
life. Bacteria routinely exchange portions of their genome; this is
their equivalent of sex. In fact, the definition of a species as true
and exclusive breeders does not even make sense for bacteria. There
are no bacterial species. The same is true for the other prokaryote
domain of life, the archaea. It was to be expected that bacteria and
archaea also might have exchanged some genes.

|

|
Figure 6.6.1 On the left, in the bacterial sexual reproduction
On the left, in the bacterial sexual reproduction
called transformation, DNA is passed from one organism to
another. Penicillin-resistant gonorrhea arose from transformation.
On the right, small circular pieces of DNA called plasmids move
independently and infect cells. They can carry genes that will make
bacteria resistant to several drugs.
|
As the tools of molecule-by-molecule genetic analysis were applied in the 1990's
(see section 5.3), it became apparent that many sections of the
genomes of eukaryotes (including plants and animals) looked a lot
like sections from bacteria and archaea. The similarity is much
greater than expected if bacteria, archaea and eukaryotes had
diverged by 3.5 Ga, as generally assumed. How could eukaryotes split
from the other domains after occurrence of multi-segmented
microfossils? Such fossils, found by UCLA paleontologist William
Schopf, presumably indicate the presence of metazoans and hence
eukaryotes. The discrepancy based on genome comparison is not subtle,
but suggests divergence more than a billion years later than
indicated by the fossil record. Should we assume that the results of
an earlier divergence has fallen victim to catastrophic extinction,
so that life history "started over", as it were, 2 Ga ago?
As more full genomes were decoded for organisms from all three domains, including our own, two
things became clear. First, we had repeatedly acquired strings of
genes by "lateral transfer" from distant taxa; microbes in particular, as evidenced by
the different ages measured by small differences in the genetic code sequences.
Second, it also became clear that advanced eukaryotes carried excess baggage in their
genome, non-working sections called introns.
(See
http://exobio.ucsd.edu/Space_Sciences/genome.htm
Bacteria
and archaea carry far less of this material; somehow they are able to
clean up their genomes. Our DNA seems to be less erasable, but our
RNA goes through some transformations in the process of coding
proteins that remove the non-coding segments.
Not only do we carry microbes in our bodies, but we depend upon some of them for
survival, as with our digestive bacteria. The ones that make us sick we call "germs",
a term with also includes viruses. Viruses are non-metabolizing bits of genetic
material, and thus, by our previous definition, are not truly "alive". They
"hijack" portions of our cellular function by inserting themselves into our
DNA ("infection"), and may permanently alter our genetic code.
|

|
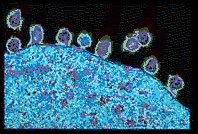
Figure 6.6.2
Human immunodeficiency virus
(HIV) particles on an infected lymphocyte.
Note the daughter HIV particles leaving to
find a new, uninfected host cell.
|
The fact that genes can cross species barriers, and that some genes insist on duplicating
themselves within the genome of a species, supports the suggestion
that genes are "selfish", that is, their apparent goal (like that of organisms) is to
survive and proliferate into all available space. In this view, the genes are the actors
and the organisms provide the background environment.
The basics of making a eukaryotic cell according to SET are as follows. The first step was
the merger of a heat and acid-loving archaeon with a swimming
bacterium like a spirochete to form the nucleocytoplasm, found in all
fungal, plant and animal cells. The conversion of free-swimming
spirochetes to cell-propelling undulipodia, cilia and so on is one of
the more controversial aspects of SET. The other steps seem to have
been confirmed by study of cell component genomes. The combined cell
was the first swimming organism with a nucleus, a protist; neither
plant nor animal nor fungus. It lived without oxygen, and was in fact
poisoned by it. The nucleus arose not from engulfing another
organism, but from the symbiotic merger of the first two organisms.
There are no free-living bacteria that look much like bare nuclei.
The next step the combined organism had to achieve was the development of mitosis, the division
of cell components during replication. The next symbiotic acquisition
was the capture of an oxygen-breathing bacterium. Next came the
ability to surround food material and create an interior space, a
vacuole, where the food might be consumed at leisure. This was
obviously a big improvement over the previous technique of nuzzling
up to food, secreting digestive fluids and re-absorbing them. The
first organism using internal digestion first appeared around 2 Ga,
that is, 2,000 million years ago.
The final component was incorporated, presumably, when food became a partner. The now
three-part cell efficiently engulfed and digested green
photosynthetic bacteria. Some green bacteria lingered on and
photosynthesized within their host, before being digested. Some hosts
learned to hold back the digestive process, to give the bacteria more
time to do their thing. A partnership was born, and eventually the
green bacteria became chloroplasts. The merged cell now was a green
micro-alga. Eventually it became the ancestor to the familiar
macro-algae and plants.
Several other classes of membrane-bounded organelles occurring in the cytoplasm between the
nucleus wall and the outer cell wall resemble bacteria in their
behavior and metabolism. Examples are plastids, mitochrondria, and
microtubules. Further study will reveal the ancestry of these highly
modified colonies of symbionts. We are their descendants.
|
|
|